BEE BREEDING
W. J. NOLAN, Apiculturist, Division of Bee Culture, Bureau of Entomology and Plant Quarantine
[ABSTRACT.] SEVERAL desirable characters have been commonly recognized among the varieties of honeybees used in the apiary, but the breeder during past years has been able to do little in combining these in one bee because in nature queen and drone mate only on the wing. These characters have to do with disease resistance, color of comb cappings, size of body parts, constancy of color markings, disposition, and swarming propensity. Because of recent work that shows the feasibility of accomplishing the insemination of queen bees by laboratory methods, the breeder is now in a position, however, to utilize the characters already available in the germ plasm of bees and to start at once toward the goal of breeding bees better adapted to present agricultural demands. It would be a great and perhaps not impossible achievement to breed a bee with the long tongue of the Caucasian, capable of reaching sources of nectar not now available to most honeybees; the gentleness of the Caucasian and the Carniolan; the white comb capping made by the black bee and the Carniolan; the reduced swarming tendency, and the resistance to common hive enemies and European foulbrood, of the Italian; the uniform marking of the Cyprian; and the industriousness of the Italian or Cyprian.
THE economic importance of bee breeding, measured in dollars, is probably greater than is realized even by the beekeeper. The potential stake of agriculture in the United States in queens alone is over $2,000,000, if a value of 50 cents, the minimum price set by the marketing agreement governing the sale of queens, is placed on each of the queens in the more than 4,000,000 colonies of bees in this country. The amount invested in bees, hive and honey-house equipment, and
the like is $35,000,000 to $50,000,000 on the conservative basis of $8
to $12 per colony. Whether a fair return is being received by the
beekeeper on these sums, plus the amount involved for his time and
labor, depends primarily on the blood represented by the queen and
by the drone with which she mates (fig. 1).
 Figure 1.—The three inhabitants of the hive: A, The worker, a female with arrested development of the egg-laying organs; B, the queen, a female with completely developed egg-laying organs but with undeveloped secondary sexual characters such as those possessed by the worker for secreting larval food, for gathering nectar and pollen, for secreting wax, and the like; C, the drone, the male bee. During the height of the season, a bee colony may consist of several thousand workers and a few hundred drones, but it normally contains only one queen at any time.
Figure 1.—The three inhabitants of the hive: A, The worker, a female with arrested development of the egg-laying organs; B, the queen, a female with completely developed egg-laying organs but with undeveloped secondary sexual characters such as those possessed by the worker for secreting larval food, for gathering nectar and pollen, for secreting wax, and the like; C, the drone, the male bee. During the height of the season, a bee colony may consist of several thousand workers and a few hundred drones, but it normally contains only one queen at any time.The public is also directly affected by the breeding problem, not merely from the standpoint of being insured an abundant supply of honey, but from the more important standpoint of having bees that will function as efficiently as possible in the pollination of many of our crops. For example, clover crops, which are so fundamental in
the dairy and livestock industries, apples and other fruits, and truck crops in greenhouses are now more or less wholly dependent on the
efforts of the honeybee. There is interest also in increasing the supply
of native beeswax, which at present comes so far short of meeting domestic demands that large quantities are imported annually. The
bee is far more valuable to American agriculture through all these other activities than through its honey production. As a matter of
fact, the activities of the honeybee are so varied and so useful that practically every person benefits by them in some way or other, and so will benefit, indirectly at least, by any improvements that result from scientific breeding.
FACTORS LIMITING PROGRESS OF THE BEE BREEDER
IN SPITE of its importance, bee breeding is only in its infancy, one reason being that promising methods for mating queens and drones under laboratory conditions had not been worked out until within the last few years. Prior to the development in 1923 of the Quinn-Laidlaw technique—discussed later in this article—whereby, through the agency of the operator, the drone organ is everted in proper position in relation to the queen for insemination to take place, and in 1926 of the Watson technique for the artificial insemination of queen bees through the transfer of sperm from drone to queen by means of a microsyringe (fig. 2), the bee breeder was forced to trust to luck in
his work, because he had to rely on chance natural matings in midair,
since in nature the honeybee mates only while on the wing away from
the hive. A successful method for obtaining natural matings of
queen and drone in confinement still remains to be developed.
 Figure 2.—L. R. Watson, who developed the Watson method for the artificial insemination of queen bees, seated at his instruments.
Figure 2.—L. R. Watson, who developed the Watson method for the artificial insemination of queen bees, seated at his instruments.Another reason for the backwardness of bee breeding is the fact that only since the application of biometric methods in studying the body parts of individual bees, a development of recent years, has progress been made in working out a method whereby the breeder can readily identify the race or strain with which he is working. Mendel, the father of modern genetics, lacked a means of distinguishing between strains when he endeavored to work out problems of bee genetics in addition to those of the genetics of peas, which brought him fame (9, pp. 142-157).
The first investigator to make a comprehensive biometric study of the honeybee was Kozhevnikov (12), who 1n 1900 endeavored to separate the various races on the
basis of physical measurements made with the aid of a microscope. He included such measurements as hooks on the wing, tongue length, length of leg segments, length of abdominal segments, length of wings, and the like. Since that time Russian investigators have been particularly active in this field.
However, the problem of checking differences between various crosses, strains, and races of the honeybee on the basis of the physical characteristics of individual bees, or even of the productivity of colonies, and then utilizing the results in breeding, is far more complicated than with other farm animals. This is due not only to the small size of the honeybee, but also to the fact that honeybee productivity represents the collective efforts of the thousands of individuals making up a colony. It is even hard to define a standard colony for purposes of any comparative experiments, since, even under the same natural conditions outside the hive, within the hive the physiological ages of the individual bees differ. Furthermore, the honeybee cannot carry on its life functions indefinitely in confinement, as can cattle, poultry, rabbits, or even pigeons.
Advances in scientific bee breeding are also limited by the variety of
demands on the breeder. Queens must be reared and tested; drones must be reared; mating of queen and drone must be accomplished; progeny must be tested for colony characteristics and production, as well as for physical characteristics of individuals; and results must be
evaluated. The scientific breeder should possess a working knowledge of the biology of the honeybee, including an intimate knowledge of its various races and strains, and of genetics, biometrics, and supplementary fields as well. In short, he should be a combination beekeeper, laboratory technician, biometrician, and geneticist, or know
how and where to utilize the services of specialists in the various fields
involved.
Another situation faced by the bee breeder is the fact that worker bees cannot be bred, and the queen and drones, mating but once in a lifetime, cannot be used in line breeding involving parent-offspring crosses. Furthermore, since queen and drone function only in reproduction, progeny tests—in other words, the collective colony
behavior exhibited by the worker progeny—must be relied upon exclusively in testing the inheritance of the queen and drone for such economically important characters as honey gathering, wax secretion, pollinating activity, and the like. Even when a parent queen is bred pure for the same color factors as the drone with which she mates, her color pattern may differ from that of her worker daughters, since in some races, the Italian being one, the causes that lead to the development of worker structures in a larva, with a resultant arrest in the development of its reproductive organs, also lead to a modification of its color pattern.
MORE KNOWLEDGE REGARDING REPRODUCTIVE PROCESS NEEDED
THE advantageous employment of present mating methods, or their
improvement, whether natural or artificial insemination is involved,
will be facilitated when the physiological processes preceding and
accompanying mating are better understood, since much remains to be known regarding the conditions most favorable for mating. For example, the consensus of opinion is that the queen mates when about a week old or shortly thereafter, but a queen 56 days old has been artificially inseminated with success. A knowledge of the external signs, if there are any, that queen and drones are ready for mating should go far in increasing the usefulness of methods of artificial insemination and also be of advantage in work involving natural in-
semination.
As a matter of fact, until recent years little was known about reproduction in the honeybee. Knowledge that drones are produced from
unfertilized eggs (parthenogenesis), at least in European varieties of
the common honeybee, dates only from the middle of the last century,
and the origin of queen and worker and the fact that the queen mates
on the wing outside the hive has been known for only about 150 years.
More has been done in studying the mating processes in the drone
than in the queen. The most complete study on the drone is that by
Bishop (4), who found that usually the drone is not sexually mature
until the ninth to twelfth day after emergence, but that some drones
are apparently incapable of mating at any age. Bishop also found the
function of the mucus, which the male organ contains in addition to sperm. During mating the sperm is merely transferred from the drone to a temporary storage place in the vagina and oviducts of the queen. The spermatozoa collect within the spermatheca during the
next few hours and may live there for 6 years or more. While still
in the oviducts and vagina, however, the sperm might all escape
through the genital opening were not this opening filled with the
mucus from the drone when the last of the sperm has entered. This
mucus quickly hardens on exposure to air, and thus forms a temporary stopper in the genital opening.
The anatomy and the physiological responses of the queen in relation to mating are being studied by Harry Laidlaw, formerly in the
employ of the Department but now a graduate student at the University of Wisconsin. This study includes the structure and functioning of the queen’s genital organs, a subject on which little is known, but a knowledge of which is demanded for the further successful development of methods, not only of artificial insemination but
even for obtaining natural matings in confinement. Laidlaw has found that a membranous fold arising from the floor of the vagina
fits so snugly in the genital passage as to be able to thwart attempts to inseminate queens artificially by his method or perhaps sometimes
to defeat efforts to insert the glass syringe by the Watson method.
MATING STATIONS AN EARLY STEP IN THE BREEDING PROGRAM
IN spite of many handicaps, the bee breeder has not been idle. During the period when he was attempting to bring the mating of the honeybee under laboratory control, he made such use of natural matings as he could by establishing mating stations in isolated bee-free localities, taking virgins and drones of desired stock there to mate. The possibility, however, that the queen may mate with a stray drone, even in a locality thought to be well isolated from local bees, reduces the value of mating stations in a scientific breeding program.
The first reported use of isolated localities in an endeavor to obtain pure matings of bees was that by Baldenstein (2) in Germany in 1848. Breeders in a number of European countries are now carrying out investigations by means of bee-mating stations. In Switzerland the
association of German-speaking beeckeepers has for a number of decades conducted a well-supported movement, inaugurated by the Swiss beekeeping leader Kramer, to maintain and improve the native
Swiss bee by having selected queens mate in isolated localities at stations stocked with drones of desired strains. The mating stations are in charge of personnel from the beekeepers’ association. Beekeepers bring virgin queens or queen cells in mating nuclei to these
stations and take back the mated queens. The method resembles the use of community-owned sires in cattle breeding.
On this side of the Atlantic such stations have not been used so much in scientific investigations. The notable exceptions are the breeding experiments conducted a number of years ago in Texas by Newell (21), and those by Sladen (34), of Canada, in progress on an island in Lake Ontario at the time of his death in 1924. Even commercial queen rearers have made little use of mating stations, because in this country isolated locations free from wild swarms of colonies of neighboring beekeepers are not easily accessible to most queen rearers. In Colorado, however, a strain of the Caucasian race was
maintained for at least 25 years through natural matings in an isolated
locality (1, p. 40). Certain commercial queen rearers in this country
have attempted insofar as possible to duplicate conditions found in
an ideal mating station by endeavoring to rid the territory in the
immediate vicinity of their mating yards of undesirable colonies of
bees. Their methods have consisted of buying up colonies found in
the neighborhood, requeening colonies nearby with desired stock
free of charge, paying rewards for wild swarms, and similar measures.
OBJECTIVES OF THE BEE BREEDER
WHAT are the immediate objectives of the bee breeder? He may
attempt to standardize the various strains now existing that are of
economic importance. He may even attempt to develop strains
that are more gentle, more disease-resistant, capable of carrying
larger honey loads, or capable of flying longer distances than strains
now available. He may attempt to develop strains with tongues
long enough to secure nectar from floral sources not now available
to the honeybee. For some regions it would seem desirable to develop
strains that fly at lower temperatures than do bees now commonly
found in the United States. This is of particular importance to
those interested in bees as pollenizing agents.
The breeder need not await the occurrence of mutations, or an
exhaustive survey of existing strains, to find some definite character
of economic importance with which to begin work. On the contrary,
he is able to start at once toward his goal, a bee better adapted to
present agricultural demands, since not one but several desirable characters have already been identified among the varieties of honeybee now used in the apiary. Other desirable characters may exist, while still others are to be expected from future mutations occurring
either naturally or under experimental conditions.
What are the characters in the available germ plasm with which
work may be begun immediately? They are related to disease
resistance, color of comb cappings, size of body parts, constancy of
color markings, disposition, swarming propensity, and the like. For
instance, the breeding of a bee in which are incorporated the long
tongue of the Caucasian, the gentleness of the Caucasian and the
Carniolan, the white capping made by the common black bee and
the Carniolan, the reduced swarming tendency and resistance to
common hive enemies and European foulbrood of the Italian, the
uniformity of marking of the Cyprian, and the industriousness of
the Italian or Cyprian would be a big achievement. Some commonly
accepted worker characters, desirable and undesirable, and the races
in which they are found are given below:
| Character | Race |
|---|
| Resistance to European foulbrood | Italian |
| Fighting wax moth | Italian, Cyprian |
| Hive cleanliness | |
| Clinging to frame during manipulation | Italian, Cyprian |
| Running on frame during manipulation | Common black, European brown |
| White cappings on honey | Common black, European brown, Carniolan |
| Water-soaked cappings | Cyprian |
| |
| Tongue length over 6.75 mm | Caucasian |
| Tongue length under 6.25 mm | Common black, European brown |
| Long legs | Caucasian |
| Uniformity of markings | Cyprian |
| Yellow scutellum |
| Gentleness | Caucasian, Carniolan |
| Viciousness | Cyprian |
| Reduced swarming tendency | Italian |
| Excessive swarming | Common black |
| Propolizing tendency | Caucasian |
BEE BREEDING IN THE UNITED STATES
INVESTIGATIONS OF THE DEPARTMENT OF AGRICULTURE
THE Department of Agriculture was a pioneer in providing the
American beekeeper with new breeding stock. It began this work by
bringing in bees from Europe in an effort to have better blood than that represented by the common black bee, introduced about the
middle of the seventeenth century, and as a result is credited with
having established the Italian race in the United States through
importations made in 1860.
In 1905 the Department sent a specialist around the world, primarily to secure the introduction of Apis dorsata Fab., the largest honeybee known, but the effort was unsuccessful. At that time the Department was distributing queens of the Cyprian, the Carniolan, and, more especially, the Caucasian race. During the last few years it has given further attention to a study of various races. As a result of this work and observations made by beekeepers in general, the recommendations of the Department as to an all-around bee for this country, for anyone other than the bee specialist or fancier, are usually limited to the Italian or Caucasian race.
The Department also led the way in this country in attempts to
develop methods for the artificial insemination of queen bees. As early as 1885, the date when it employed its first full-time specialist in beekeeping, it set forth as one of his duties: “To make experiments in the methods of artificial fertilization and, if possible, demonstrate the best process by which the same may be accomplished.” Some slight success was reported from this early venture, and a method described in which a fine-pointed syringe was employed, as in the present Watson method, but this early work was not followed up. In 1907 the Department conducted unsuccessful attempts to obtain the mating of queens on the wing within enclosures.
Within the last few years the Department (23) has greatly simplified the technique and equipment for the Watson method, reducing
the cost of equipment and making it possible to operate on several queens more or less simultaneously, thus increasing the output (fig. 3).
Some of the special features of this new equipment differentiating it
from that developed by Watson are as follows: Only one microscope is needed in using a large number of syringes; a cheap-and easily made
manipulator is provided for the microsyringe; the queen is held in a glass tube with only her abdominal tips projecting, instead of being
bound by thread to a small wooden block hollowed out to fit her body;
and the queen’s abdominal tips are spread apart by a pair of metal hooks held mechanically instead of by forceps held by hand (fig. 4). With this equipment the genital opening of the queen can be found quickly under the microscope, and the microsyringe then can readily
be brought into correct position for discharging the sperm without
injury to the queen.

 Figure 3.—Multiple set-up devised in the Department of Agriculture for inseminating several queens more or less simultaneously. The set-up shown consists of 6 separate units, each equipped with a microsyringe and apparatus to hold the queen during the operation. One microscope serves the entire set-up, being moved from unit to unit as needed.
Figure 3.—Multiple set-up devised in the Department of Agriculture for inseminating several queens more or less simultaneously. The set-up shown consists of 6 separate units, each equipped with a microsyringe and apparatus to hold the queen during the operation. One microscope serves the entire set-up, being moved from unit to unit as needed.

Figure 4.—Separate unit of the multiple set-up, showing queen and microsyringe in position. The microsyringe differs somewhat in construction from the original Watson type but works on the same principle. A less fragile microsyringe, all metal except for the glass point and with the movement of the plunger in and out caused by direct action, has since been devised in the Department.A durable microsyringe, all metal except for the minute plunger
tube of glass that holds the sperm, has been developed from an automatic pencil by substituting a fine wire plunger for the lead and attaching the plunger tube to the point of the pencil. The earlier syringe consisted of a piece of glass tubing drawn out to proper shape for
holding the plunger and plunger mechanism, as well as for attaching
the plunger tube.
Sperm to fill the syringe is obtained by causing partial eversion of
the drone organ by pressure on the drone, after which the bulb containing both sperm and mucus is pulled loose with a pair of forceps. With the bulb still held in the forceps, the end containing the white mucus is cut away and the remainder, forming a minute sac containing the cream-colored sperm, is slipped over the point of the syringe. The sperm is then drawn into the plunger tube.
The Department has also modified the Quinn-Laidlaw method for
artificial insemination, commonly referred to as a “hand-mating” method, by using a glass tube to hold the queen and a small spring
placed inside her abdominal tips to hold them apart (14). A microscope is now used during the process. As when the method was first
developed, the drone is so held by hand as to cause partial eversion
of its genital organs and is then so placed in relation to the queen that
the transfer of sperm will be accomplished after complete eversion
has taken place.
Biometric data on common races of the honeybee and on the progeny of various crosses made by controlled matings are being collected by the Department to determine racial or strain characters
of a quantitative nature and their behavior in crosses. The data collected so far consist mostly of linear measurements of various parts of the head, thorax, and abdomen. The specimens are mounted on slides under a microscope and the measurements are made on projected images (fig. 5). Those of head parts (fig. 6) include tongue and scapus (the long joint of the antenna). On the thorax they cover counts of hooks on the hind wing and dimensions of the fore wings (fig. 7, A) and hind legs (fig. 8). On the abdomen the
measurements deemed important are width of the third tergum (the upper surface of the third segment), longest width of third sternum
(lower surface of third segment) across its left wax plate, and longest dimension of the wax
plate itself. The first two abdominal segments, because of their shape and lack of wax plates, do not lend themselves to such measurements (fig. 9).
 Figure 5.—Apparatus used in the Department of Agriculture for measuring external parts of bees. A micrometer eyepiece with a movable scale is attached to the microscope.
Figure 5.—Apparatus used in the Department of Agriculture for measuring external parts of bees. A micrometer eyepiece with a movable scale is attached to the microscope.

Figure 6.—A, Head of bee, showing: a, Antenna; b, gena; c, clypeus; and d, labrum. Tongue is shown extruded but cut off. B, Extended tongue, showing: a, Submentum; b, mentum; c, ligula; and d, labial palpus. (Drawing by Snodgrass.)
 Figure 7.—A, Front wing of honeybee, showing: a-b, Proximal length measured; b—d, distal length measured; c, third cubital cell. B, Cubital cell enlarged to show method of measuring cubital index (a/b).
Figure 7.—A, Front wing of honeybee, showing: a-b, Proximal length measured; b—d, distal length measured; c, third cubital cell. B, Cubital cell enlarged to show method of measuring cubital index (a/b). (Drawing by Snodgrass.)
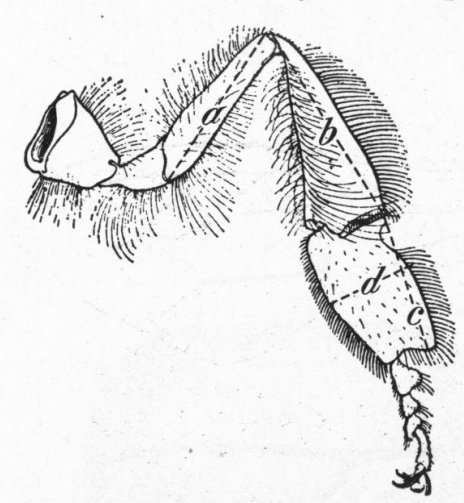 Figure 8.—Hind leg of worker bee, showing measurements made: a, Length of femur; b, length of tibia; c, length of metatarsus; d, width of metatarsus
Figure 8.—Hind leg of worker bee, showing measurements made: a, Length of femur; b, length of tibia; c, length of metatarsus; d, width of metatarsus (Drawing by Snodgrass)
 Figure 9.—Lateral view of worker abdomen showing the six abdominal segments, including a portion of the thorax at left: a, Left end of third abdominal sternum; b, left end of third abdominal tergum.
Figure 9.—Lateral view of worker abdomen showing the six abdominal segments, including a portion of the thorax at left: a, Left end of third abdominal sternum; b, left end of third abdominal tergum. (Drawing by Snodgrass.)
The size of the foregoing parts is of economic interest as well as of value in distinguishing races or strains, because nearly all of them function directly in the gathering of nectar and pollen and in the production of wax. The tongue, for instance, is of importance in nectar gathering. The wings are important in all activities taking the bees away from the hive. The hind legs carry pollen. The abdominal segments contain the wax plates, and may serve to limit the size of the honey sac.
Characters of a qualitative nature are also being studied. These include gentleness, industry, disease resistance,
and the like. At the present time the Department is engaged in a
cooperative undertaking with the States of Iowa, Texas, Wisconsin,
and Wyoming, to search for stock resistant to American foulbrood,
and then to combine the factor for resistance, if any be found, with
other desirable factors by breeding.
During the past year the Department has employed a trained
geneticist for the first time in its beekeeping work. His immediate
genetical problem is to determine whether heritable factors affect
supersedure, especially the premature supersedure of queens sold
commercially. By "supersedure” is meant the replacement of the
queen by another reared by the colony apparently for that purpose.If heritable factors play a part in this behavior, they may lie in the
behavior of the queen alone—lack of fecundity, for example—or
they may lie in the behavior of the workers—for example, undue
hostility to a queen reared in another colony.
STATE BEE-BREEDING WORK
Only a few States have been engaged in breeding work. A number of years ago Shafer (30), at the Michigan Agricultural Experiment Station, made an unsuccessful attempt to obtain controlled matings of queen bees by natural insemination. The New Jersey Station conducted bee-breeding work in an isolated location for a number of
years in connection with a study of disease resistance. More recently
the Iowa Station, in cooperation with the American Bee Journal, has been engaged in a program to find stock resistant to American
foulbrood and to breed for this character. The Texas Station is endeavoring to breed a bee that will better withstand Texas weather
and possess greater honey-gathering ability than present strains. The
College of Agriculture of the University of California is studying the
correlation between physical characters of queens and their productivity, and is also studying environmental factors that influence the development of queens. The College of Agriculture of the University of Minnesota a number of years ago conducted work on the artificial insemination of queen bees by the syringe method, with some slight success. In this work, performed by Jager and Howard (10), the sperm was diluted with salt solution.
Merrill (16), while in charge of apicultural work at the Kansas State Agricultural College, became the American pioneer in the biometric study of the activities of the colony as a whole. He attempted to correlate honey production with tongue length, size of honey sac, and other physical characters. A few years ago Munro (19), while a student at this college, made a study of color inheritance, using natural matings. At present E. J. McNay, a graduate student at the same institution, is making use of the Watson method in breeding work.
WORK BY PRIVATE BREEDERS
The outstanding bee-breeding work of a private nature is that by Watson (36, 37), who developed the method for the artificial insemination of queen bees that bears his name, and who now holds a Guggenheim fellowship for the study of bee breeding. He is making a survey of tongue lengths of various strains in an endeavor to develop a longer tongued bee than is now available. He is also trying by artificial insemination to perpetuate the "Albino" bee, a mutation of the Italian that appeared in this country about 50 years ago but has now apparently disappeared except for stock in Watson’s apiary. This bee is marked by an unusually large quantity of long gray or white hairs, especially on the abdomen.
Jay A. Smith is using the Watson method in an endeavor to breed a more yellow and gentler bee. In a private communication he reports success in diluting the sperm with a saline solution before taking it into the microsyringe.
About 25 years before the present methods for the artificial insemination of queen bees were developed, Root (26) greatly stimulated interest in bee breeding through publicity given a "red clover queen” of the Italian race, from which was bred progeny said to have exceptionally long tongues for Italian bees. This strain soon disappeared. Among present-day workers in this country who are endeavoring to breed bees by natural insemination may be mentioned Ralph Benton, and Erwin Alfonsus, the latter formerly with the University of Wisconsin.
Balinkin (3) and Clifford Muth have pioneered in this country in
the application of ultraviolet rays to young queens, but details of their
method have not yet been published. An increase in egg laying and progeny with better dispositions were
reported, but it was not claimed that this modified behavior is transmissible.
BEE BREEDING IN FOREIGN COUNTRIES
Numerous foreign workers are interested in bee breeding (see the appendix). The work of the German-speaking beekeepers’ association in Switzerland has already been mentioned. Breeding along the same lines is in progress in Germany and elsewhere in central Europe. Zander (40, p. 193), assisted by A. Himmer, for years used isolated mating stations to breed an improved strain of the European brown bee, as part of the program of the bee-research institute at Erlangen. Armbruster (2) is another German investigator who has worked in this field.
Götze (7), also of Germany, has been engaged in a biometric study
of progeny of crosses obtained at mating stations. He has reported unsatisfactory results in the use of artificial insemination. He places considerable reliance upon certain indices, which are based on proportions involving certain wing cells, as a means of identifying various strains with which he works. The most important of these is the cubital index, which is the ratio of the two parts into which the line joining the two corners of the third cubital cell of the front wing on the side bounded by the cubital vein is divided when a perpendicular is dropped from this line to the second recurrent vein (fig. 7, B, illustrates this measurement).
Alpatov (1, p. 24) of the Union of Soviet Socialist Republics, has perhaps made a wider use of biometrical indices in studying the differences of races or strains than has any other investigator. Some indices used by him are the proportion between width of the first wax gland and width of the sternum that
contains it, length of the hind metatarsus and width of third abdominal tergum, length of hind tibia and width of third abdominal tergum, length of hind femur and width of third abdominal tergum, distal length and proximal length of front wing, and width and proximal length of front wing (figs. 7, 8, and 9).
Jaroslav Rytif, of Czechoslovakia, has made observations on the inheritance of measurable characters of various strains, including “strain 47”. This strain was developed, or rather perpetuated, by the Austrian queen breeder Sklenar (31, p. 5) from a colony that had been
outstanding in his apiary. Sklenar thinks the strain is descended from
a mixture of the Carniolan and Italian races, although the workers
show no yellow color. They are usually gray but occasionally one is
reddish brown. Rytif (27) has also studied inheritance of amount of
coloration on abdominal segments, and has devised a workable system
for obtaining quantitative data on this characteristic.
A few years ago Mikhailoff, of the Union of Soviet Socialist Republics, reported considerable success with the Watson method. From the same country also came the Malyschev method (15). This differs from the Quinn-Laidlaw method in that the drone organ is
first dissected out under the microscope and then placed in position
in the queen. Prell (25), of Germany, working independently,
reported success with a method like that of Malyschev.
GEOGRAPHICAL STATUS OF DEVELOPMENT OF BREEDING TECHNIQUE
IN THE United States successful methods for the controlled mating of queen and drone under laboratory conditions have been developed and existing races of the honeybee have been tried out. In western and central Europe workers have made praiseworthy attempts at the improvement of bees by selective breeding in isolated mating stations. In the Union of Soviet Socialist Republics breeding work has centered largely around biometric studies in an endeavor to reduce the characteristics of races and strains to some sort of mathematical formula whereby each can be more easily distinguished. Some work has been carried out with artificial insemination.
TOO LITTLE is known concerning the cytology of the honeybee. For a long time the work of Nachtsheim (20), which seemed to show that the diploid number of chromosomes (that of queen and worker) is 32 and that the haploid number (that of drones) is 16, has been accepted as standard. In recent work, however, Sanderson (29) has questioned whether Nachtsheim has reported the normal numbers or whether they are not 16 and 8 for queen and drone, respectively. The characteristic shape of any of the chromosomes has not yet been determined.
Even the type of sex chromosome and the method of sex determination in the honeybee are still undetermined. Nachtsheim holds that
the sex chromosomes are of the X type, the female possessing two and the male but one, since the drone arises from an unfertilized egg. Likewise the female possesses twice as many other chromosomes (autosomes) as does the male. Consequently, according to Nachtsheim, sex is determined on a quantitative basis—doubling the sex chromosome and autosome complement results in a female instead of a male. This view is not in accord with the theory of genic balance that is now applied to so many organisms. According to this theory,
sex is not determined on a quantitative basis, but is the resultant of a tendency towards maleness or femaleness in genes of the autosomes taken as a group and an opposing tendency in genes of the sex chromosomes.
In contrast to Nachtsheim’s theory of sex chromosomes in the honeybee, Whiting (39) holds that in the parasitic wasp Habrobracon juglandis Ashmead, another hymenopterous form in which males develop parthenogenetically, the two sex chromosomes of the female are of the XY and not of the XX type. Consequently two types of haploid males develop, depending on whether they contain a single X or a single Y chromosome. Outwardly they appear alike, but they can be identified genetically. They are produced in equal numbers. Of even greater interest, two types of biparental males are to be found and these contain either two X or two Y chromosomes. Biparental males, like the females, carry two sets of autosomes, but cannot be distinguished from the ordinary parthenogenetic males unless the parent male possesses a factor that is dominant to the homologous factor carried by the homozygous mother.
Whiting (39), basing his conclusions on experimental evidence, explains sex determination in Habrobracon by assuming that the X chromosome carries the factors F.g while the allelomorphs (f.G) of these two factors are carried by the Y chromosome. The presence of both F and G, classed as "complementary factors” by Whiting, is necessary for the development of a female, otherwise only a male will result. The dominants F and G would both be present, of course, whenever fertilization of the egg represents the union of X and Y, since the egg would then carry the factors F.g/f.G. The union of a sperm and an egg each of which has an X chromosome, or each of which has a Y chromosome, could only result in a biparental male, because in either event the fertilized egg would contain only one of the two dominant complementary factors necessary if a female is to result. The factors in the one case would be F.g/F.g, and, in the other f.G/f.G. Whiting’s line of reasoning would hold, whether F and G each represents only a single dominant gene or whether they are the symbols of a group of dominant genes carried in their respective chromosomes (X and Y).
Whiting (38) first encountered biparental Habrobracon males in a
culture headed by a female homozygous for orange-colored eyes but
mated to a male with black eyes, the dominant color. Instead of all
the sons having orange eyes, a few resembled the daughters in having
black eyes. Such males usually proved sterile, although in certain
instances in which they were mated with females homozygous for
orange-colored eyes a few daughters were obtained, some with orange
eyes and some with black eyes. A knowledge of whether or not
Whiting’s results on Habrobracon hold good for the honeybee awaits
further work.
The fact that the drone is haploid seems to have led to the impression that drones from a heterozygous queen will be only of two kinds, corresponding to the queen’s maternal and paternal inheritance. Such a conclusion holds only if one pair of unit factors or linkage groups is under consideration. For example, normally half the drones from a queen heterozygous for color—say, yellow and black—would carry the factor for yellow and half of them the factor for black if these were unit factors. Her drones would fall into four types, however, if their inheritance of any two pairs of unlinked factors is considered. Thus, if it is assumed that the characters for tongue length and those for color are not linked, but that each is determined merely by a unit factor, a queen heterozygous for these two factors would produce long-tongued yellow drones, short-tongued yellow drones, long-tongued black drones, and short-tongued black drones. The greater the number of pairs of unlinked factors that are under consideration, the greater will be the number of types into which the drones can be classified.
RESULTS OBTAINED FROM BEE BREEDING
It is unfortunate that Mendel’s detailed records have never been
brought to light. His studies were not confined to one race of the
honeybee, since it is reported (9) that he worked on the heather or
Dutch bee, the Italian, the Carniolan, the Egyptian, and the Cyprian,
in addition to the native bee of his region.
Since so much of the work concerned the development of a technique for breeding the honeybee under controlled conditions, and a
biometric study to determine measurable physical characters of
individual members of a colony as well as characteristic colony
behavior marking races or strains, strictly genetic results are scanty
as yet, and often of a somewhat general nature.
The work of the Department has verified the fact that honeybees
can be successfully carried from generation to generation by means
of artificial insemination, seven successive generations having been
obtained in three seasons’ work. Allowing the queen 16 days to
develop from egg to adult, 7 days in which to mate, and 3 more in
which to begin egg laying, there should theoretically be a laying
queen of each successive generation every 26 days, but the normal
hazards and delays of beekeeping have greatly increased this period
in actual practice.
The Department has verified Watson’s feat (37) of transferring
sperm from the spermatheca of one queen to that of another. The queen from which sperm was transferred had been dead for several hours. This procedure makes possible a type of line breeding, involving crosses of a drone' with his offspring, although on an extremely
limited scale, because the supply of sperm from the original drone
cannot be replenished but will be diminished when transferred from
queen to queen.
The work of the Department also indicates that color and tongue length can be inherited independently of each other and that each race has its own size characteristics, which are inheritable and more or less constant within fairly definite limits. Judging from preliminary data, some correlation exists between tongue length and number of wing hooks, and also between tongue length and length of scapus. Götze’s statement (7, p. 227) of the correlation between tongue length and length of labial palpi also seems borne out by the work thus far.
A hitherto undescribed mutation, involving a yellow coloration on
the face of the bee—principally on the clypeus, genae, labrum, and,
at times, bases of antennae (fig. 6, A)—was found in an Italian strain
in the Department’s apiary formerly at Somerset, Md., and an attempt
is being made to obtain a homozygous strain. However, it appears
to be a recessive character.
The development of stock especially resistant to European foulbrood was announced as the result of selective breeding experiments in
New Jersey (6). In describing the first season’s work of the Iowa Agricultural Experiment Station to find stock resistant to American foulbrood, Park (24) concluded that variation in resistance to American foulbrood exists. Some evidence of the inheritance of number of egg tubules has been reported in a private communication from J. E. Eckert, describing work at the University of California.
Since the World War an independent breeder, Henry Brown of
New Jersey, has developed a light-colored strain, reputed to be very
gentle, by crossing the Cyprian and Italian races, according to a
private communication.
Even years before Brown’s work the “golden" Italian bee, a strain with the anterior portion of each of the first five abdominal tergites marked with a golden band, instead of only each of the first three as in the ordinary Italian, was developed in this country, supposedly
by crossing the Italian with the Cyprian. This golden Italian, while
held to be gentle and a beautiful bee, had little repute as a honey
gatherer. Sladen (32) developed a “British golden” by crossing the
English strain of the common black bee, the Italian, and the “American
golden.” By continued selective breeding in an isolated locality, a
bee was obtained which Sladen claimed combined the hardiness and
industriousness of the black with the prolificness of the Italian,
although its area of golden coloration was not so extensive as that of
the "American golden.” No attempt was made to increase the area
of golden coloration, however, since this character was used merely
to provide breeding stock with drones of such a color that their progeny
could easily be distinguished from that of the common black drone,
which prevailed elsewhere in the locality.
Jay A. Smith reports in a private communication that in crosses
obtained by the Watson method no correlation seemed to exist
between color and gentleness.
The reports of the work in Switzerland are of especial interest to
beekeepers, since rather extensive records are kept showing the returns from improved (veredelte) stock, meaning that developed at
the mating stations, as compared with those from ordinary or unimproved stock. Thus from 1915 to 1928, according to a survey of
certain apiaries which contained colonies of both types of stock, the honey crop of the improved stock averaged larger per colony each year than that of the unimproved stock, ranging from 20 percent more in 1923 to 100 percent more in 1924 (13, p. 30).
Mikhailoff (18) used the Watson method of insemination to demonstrate that the spermatozoa from white-eyed drones are active, and he succeeded in obtaining white-eyed workers and queens. White-eyed drones apparently do not see and do not mate in nature. That the gene for white eyes is recessive to the normal eye color is shown by the fact that with natural mating a queen laying eggs from which white-eyed drones arise gives only workers with normal eye color.
In the Department apiary formerly at Somerset, Md., half of the
drones from an Italian queen that produced white-eyed drones were
white-eyed and half were black-eyed, a proportion also observed by
other investigators. A few of the white-eyed drones were seen to
fly off into the air, but none was seen to reenter the hive. Some, on
leaving the hives, made short hops or flew in small circles away from
the hive entrance. In the fall only black-eyed drones were found in
the hive with this queen. Whether at this season the workers
removed the white-eyed drones before emergence is problematical, since no white-eyed drone pupae were found in any of the sealed cells. A number of virgins were reared from this queen, but attempts to inseminate them with sperm from the white-eyed drones were unsuccessful. One of these virgins inseminated with sperm from a normal
drone, however, produced some white-eyed drones. Part of the worker progeny of the original queen were a lighter yellow on their
abdomens than the ordinary Italian bee.
As for the inheritance of quantitative characters, Mikhailoff reported (17) that a Caucasian queen from a colony with a tongue
length of 6.9393 mm was mated with a central-Russian black bee with
a tongue length of 6.1729 mm, and the resulting cross gave a bee with
tongue length of 6.705 mm. Twelve percent of these hybrids had
tongue lengths of 7.0282 mm, this being greater than that of either
parent race. Two Caucasian queens that were inbred with "brother”
drones had progeny with shorter tongues than those of their worker
sisters. Other quantitative data are included and, although they
represent a very scanty number of colonies, they are of interest
because they are the first statistical data to be published that are
derived from controlled crossing of the honeybee.
Alpatov (1) has concluded as a result of his studies in the Union of Soviet Socialist Republics that, in the area covered, absolute size of body and of wings, relative size of the wax gland, and the color of the abdomen seem linked. The same is true concerning length of tongue and length of hind legs. He (I, p. 40) has pointed out that the constancy of color of the yellow and the black races in the United States shows that color is an inherited racial characteristic.
Watson (37) has stated that the albino bee is dominant over the common black bee.
Newell (21) crossed yellow Italians and black Carniolans and found
yellow dominant. Watson (37) crossed a black virgin and a yellow
drone and obtained all black offspring. It may be assumed that Watson used the common black or Dutch bee. Sladen (33, p. 64),
in crossing a golden queen with pure black drones, obtained “intermediate" workers that had black bands on the posterior margins of
the second to fifth abdominal tergites, increasing in width from tergite to tergite toward the rear. The sixth tergite was black. As a matter of fact, the last abdominal tergite is black in all varieties of the European honeybee, no mutation showing a different color having
yet been reported.
In work at the Department (22) a daughter of a European brown
bee crossed with a yellow drone produced workers that might have
been taken casually as a dark strain of three-banded Italians, possibly
because of the blackness of the posterior bands of each of the first
three abdominal tergites. Practically the same results were obtained
later with Caucasians and Italians.
Mikhailoff (17), in crossing queens of the central-Russian black
bee with golden Italian drones, found yellow to be the dominant color
in 90 percent of the progeny.
Götze (8) holds that the yellow scutellum, such as occurs in the
Cyprian and other eastern races, is the expression of a dominant unit
factor. He found that a golden queen mated to a black drone gave
workers with yellow scutella. These workers, however, as regards
abdominal coloration, were not golden, but had abdominal segments
with decided black edges, as in the work by Sladen and the writer. Götze (8, p. 71) therefore calls the F1 an “intermediate form" and further assumes that a separate gene is responsible for the black edges on the abdominal segments.
Götze has found in various colonies both black and brown drones
as well as some termed by him "leather-colored.” To account for
these various colorations he assumes a basic color factor, N, which
requires the presence of an activating factor to give rise to color. He
assumes that not one but two activating factors or their allelomorphs
are carried. These are M, a factor for black, and B, a factor for brown. B is inhibited by M. When only their allelomorphs bm are present,
leather-colored individuals arise. Götze states that m is a factor
for leather-colored but does not give the nature of b. A black drone,
according to this scheme, would have the genic composition NBM or NbM; a brown drone, NBm; and a leather-colored drone, Nbm. In addition, Götze found that certain eggs did not hatch, and he holds that they had the genic combination nbm, which he assumes is lethal. In an actual count, omitting the supposed lethal cases, he found 110 black, 48 brown, and 21 leather-colored workers.
Stucki (35) has suggested the possibility that certain definite excesses or deficiencies from the normal wing venation may be characteristic of certain strains of Swiss bees, although he postpones definite conclusions until more material is accumulated. Like Casteel and Phillips (5) and others, he found drones especially variable. Rytit (28) reports, as a result of crossing the Iskra strain of bees from Czechoslovakia with the Nigra strain of the European brown bee—
the latter strain having a lower cubital index (fig. 7, B)—that the
lower index is completely dominant over the higher.
The dominance or recessiveness of certain characters of the honeybee
that have been reported as heritable is shown below. It is to be
remembered that most of the data need further confirmation before
being accepted and that, while many of the characters listed seemingly
have little or no direct economic importance in themselves, yet, if
they happen to be linked with characters of economic importance,
perhaps they may ultimately prove of use as an index to the presence
of such characters.
Dominance or recessiveness as shown in F1 generation
| Dominant factors | Recessive factors | Reported by— |
|---|
| Eye color normal | Eye color white | Mikhailoff (18) |
Abdominal marking:
Italian | Abdominal marking:
European brown | Mikhailoff (17) |
| Carniolan | Newell (21) |
| Common black | Italian | Watson (37, p. 40) |
| Albino | Common black |
| Cubital index low | Cubital index high | Rytiř (27) |
| Scutellum yellow | Scutellum black | Munro (19) |
| Clypeus, genae, and labrum dark | Clypeus, genae, and labrum yellow | Nolan (this article, p. 1534 [which does not exist -ASC]) |
| Use of wax for sealing crevices, etc. (Carniolan) | Use of propolis for sealing crevices, etc. | Newell (21) |
| | | |
Some crosses in which dominance or recessiveness is not shown for certain characters by F1 generation
| Cross | Character | Reported by— |
|---|
| European brown X Golden | Abdominal coloration | Götze (8, p. 71) |
| Common black X Golden | Sladen (33, p. 64) |
| European brown X Italian | Nolan (22) |
| Long X short | Tongue length | Götze (8, p. 73) |
Some linked characters
| Characters | Reported by— |
|---|
| Absolute size of body and wings, relative size of wax glands, color of abdomen | Alpatov (1, p. 48) |
| Tongue length, labial palpi | Götze (8, p. 227) |
| Tongue length, number of egg tubules | Komarov and Alpatov (11) |
| Yellow color, light weight (drones) | Munro (19) |
| Black color, heavy weight (drones) |
Now that a notable advance in the technique of accomplishing matings has been made, methods for obtaining biometric data have been worked out, and germ plasm that possesses a number of desirable characters is available, the way appears open for a worth-while advance in bee breeding, although quick results should not be expected.
(1) Alpatov, W. W. 1929. BIOMETRICAL STUDIES ON VARIATION AND RACES OF THE HONEY BEE (APIS MELLIFERA L.). Quart. Rev. Biol. 4: 1-58, illus.
(2) ARMBRUSTER, L. 1919. BIENENZUCHTUNGSKUNDE. VERSUCH DER ANWENDUNG WISSENSCHAFTLICHER VERERBUNGSLEHREN AUF DIE ZUCHTUNG EINES NUTZTIERES. 120 pp., illus. Leipzig, Berlin.
(3) BaLINKIN, I. 1930. THE WONDERS WITHIN THE SPECTRUM. Beekeepers’ Item 14:193-196.
(4) Bishop, G. H. 1920. FERTILIZATION IN THE HONEY-BEE. I. THE MALE SEXUAL ORGANS: THEIR HISTOLOGICAL STRUCTURE AND PHYSIOLOGICAL FUNCTIONING. II. DISPOSAL OF THE SEXUAL FLUIDS IN THE ORGANS OF THE FEMALE. Jour. Expt. Zool. 31: 225-[226], 267-286, illus.
(5) Casteel, D. B., and Phillips, E. F. 1903. COMPARATIVE VARIABILITY OF DRONES AND WORKERS OF THE HONEY BEE. Biol. Bull. 6: 18-37.
(6) FilMeR, R. S. 1931. BEE INVESTIGATIONs. N. J. State Agr. Expt. Sta. Rept. 52:228-238.
(7) Götze, G. 1930. VARIABILITÄTS UND ZÜCHTUNGSSTUDIEN AN DER HONIGBIENE MIT BESONDERER BERÜCKSICHTIGUNG DER LANGRÜSSELIGKEIT. Arch. Bienenk. 11: 185-279.
(8) ―1936. TECHNIK UND ERGEBNISSE VON ERBVERSUCHEN MIT DER HONIGBIENE. Deut. Imkerführer 10: 68-74.
(9) Iltis, H. 1924. GREGOR JOHANN MENDEL, LEBEN, WERK UND WIRKUNG, 426 pp., illus. Berlin.
(10) Jager, F., and Howard, C. W. 1914. THE ARTIFICIAL FERTILIZATION OF QUEEN BEES. Science (n. s.) 40: 720.
(11) Komarov, P. M., and ALpaTov, W. W. 1935. BEITRAGE ZUR KENNTNIS DER VARIABILITAT DER HONIGBIENE. Arch. Bienenk. 15: 11-20.
(12) KozheEvNIKOV, G. A. 1900. CONTRIBUTIONS TO THE NATURAL HISTORY OF THE BEE (APIS MELLIFERA L.). Pt. 1, illus. Moscow. [In Russian.]
(13) Kramer, U. 1930. DIE RASSENZUCHT DER SCHWEIZER IMKER. Aufl. 7, revised by M. Jiistrich. 208 pp., illus. Aarau.
(14) Laidlaw, H. J., Jr. 1932. HAND MATING OF QUEENBEES. Amer. Bee Jour. 72: 286.
(15) MalyscHEV, S. I. 1924. [ARTIFICIAL INSEMINATION OF THE HONEYBEE.] Chelovek i Priroda 1924: 579-586. [In Russian. Original not seen.}
(16) MerriLL, J. H. 1922. THE CORRELATION BETWEEN SOME PHYSICAL CHARACTERS OF THE BEE AND ITS HONEY-STORING ABILITIES. Jour. Econ. Ent. 15:125-129. ¥
(17) MikHAILOFF, A. S. 1930. [ON THE INHERITANCE OF THE COLOR AND SOME QUANTITATIVE CHARACTERS IN THE F; IN THE HONEYBEE.] Opitnaia Paseka 1930: 215-228, illus. [In Russian.]
(18) ―1931. UBER DIE VERERBUNG DER WEISSAUGIGKEIT BEI DER HONIGBIENE (APIS MELLIFERA). Ztschr. Induktive Abstam. u. Vererbungslehre 59: 190-202.
(19) Munro, J. A. s 1925. INHERITANCE IN THE HONEYBEE. Amer. Bee Jour. 65: 337-338.
(20) NachtshEIM, H. 1913. CYTOLOGISCHE STUDIEN UBER DIE GESCHLECHTSBESTIMMUNG BEI DER HONIGBIENE (APIS MELLIFICA L.). Arch. fiir Zellforsch. 11: 169-241, illus.
(21) NeweLL, W. 1915. INHERITANCE IN THE HONEY BEE. Science (n. s.) 41: 218-219.
(22) Nolan, W. J. 1929. SUCCESS IN THE ARTIFICIAL INSEMINATION OF QUEEN BEES AT THE BEE CULTURE LABORATORY. Jour. Econ. Ent. 22: 544-551.
(23) ―1932. BREEDING THE HONEYBEE UNDER CONTROLLED CONDITIONs. U. S. Dept. Agr. Tech. Bull. 326, 49 pp., illus.
(24) Park, O. W. 1936. DISEASE RESISTANCE AND AMERICAN FOULBROOD. Amer. Bee Jour. 6: 12-15.
(25) PreLL, H. 1927. DIE KUNSTLICHE BEFRUCHTUNG DER BIENENKONIGIN. Leipziger Bienen-Ztg. 42: 225-230; also in Mairkische Bienen-Ztg. 17:236-241; and 10th Cong. Internatl. Zool., 1927, 10 (pt. 1):[455]-462.
(26) Roor, E. R. 1900. OUR $200 RED CLOVER QUEEN AND HER DAUGHTER. Gleanings Bee Cult. 28: 813.
(27) Rytiř, J. 1929. Nová METODA VYŠETROVÁNÍ BARVY VČEL. Věela Moravská 52:260-264.
(28) ―1936. DĚDIČNOST LOKETNIHO INDEXU MÍŠENCŮ NIGRA x ISKRA. Česky Věelar 70: 13-16.
(29) SanDERsON, A. R. 1932. THE CYTOLOGY OF PARTHENOGENESIS IN TENTHREDINIDAE. Genetica 14: 321-501, illus.
(30) SHAFER, G. D. 1917. A STUDY OF THE FACTORS WHICH GOVERN MATING IN THE HONEY BEE. Mich. Agr. Expt. Sta. Tech. Bull. 34, 19 pp., illus.
(31) SKLENAR, G. 1934. STAMM „47" UND SEINE zUCHT. 159 pp., illus. Mistelbach, Austria.
(32) Sladen, F. W.L. 1909. BREEDING THE BRITISH GOLDEN BEE IN RIPPLE COURT APIARY. Brit. Bee Jour. 37: 482-486; 497-499; 507-509; 511-512.
(33) ―1913. QUEEN-REARING IN ENGLAND. WITH NOTES ON A SCENT-PRODUCING ORGAN IN THE WORKER-BEE; AND HOW POISON IS COLLECTED BY THE HONEY-BEE AND BUMBLE-BEE. Ed. 2, 86 pp., illus. London.
(34) ―1920. QUEEN-MATING EXPERIMENTS. Gleanings in Bee Cult. 48: 717-718.
(35) Stucki, H. 1936. HAT UNS DAS ADERBILD DES BIENENFLUGELS WICHTIGES ZU SAGEN? Schweiz. Bienen-Ztg. 59: 37-40.
(36) Watson, L. R. 1927. CONTROLLED MATING OF QUEENBEES. 50 pp., illus. Hamilton, III.
(37) ―1927. CONTROLLED MATING IN THE HONEYBEE. Iowa State Apiarist Rept. 1927: 36-41.
(38) WHITING, P. W. 1921. HEREDITY IN WASPS. A STUDY OF HEREDITY IN A PARTHENOGENETIC INSECT, THE PARASITIC WASP, HABROBRACON. Jour. Heredity 12: 262-266.
(39) ―1935. SEX DETERMINATION IN BEES AND wasps. Jour. Heredity 26:263-278.
(40) ZANDER, E. 1930. DIE ZUCHT DER BIENE. Ed. 3, 310 pp., illus. Stuttgart.
APPENDIX
Some Workers Identified with Bee Improvement at State and Federal Experiment Stations and Other Institutions
California, Davis: J. E. Eckert.
Connecticut, New Haven: L. C. Curtis.
Iowa, Ames: O. W. Park.
Kansas, Manhattan: E. J. McNay.
New Jersey, New Brunswick: R. S. Filmer.
Texas, San Antonio: H. B. Parks.
Wisconsin, Madison: H. Laidlaw.
Other institutions:
Brigham Young University, Provo, Utah: J. Fleming Wakefield.
Alfred University, Alfred, N. Y.: L. R. Watson.
Depauw University, Greencastle, Ind. Jay A. Smith.
United States Department of Agriculture, Bureau of Entomology and Plant Quarantine:
Baton Rouge, La.: Otto Mackensen.
Beltsville, Md.: W. J. Nolan.
Laramie, Wyo. A. P. Sturtevant
Some Workers in Foreign Countries Identified with Bee Improvement
Austria, Mistelbach: Guido Sklenar.
China, Fukien Christian University, Foochow: C. R. Kellogg
Canada, Dominion Experimental Farms, Ottawa: C. B. Gooderham
Czechoslovakia, Krpy: Jaroslav Rytiř
Czechoslovakia, Ober-Hohenelbe: Egon Rotter
Egypt, Matarich, Cairo: A. Z. Abushady
Germany, Landesanstalt für Bienenzucht, Erlangen: E. Zander and A. Himmer.
Germany, Landwirtschaftliche Hochschule, Hohenheim bei Stuttgart: G. A. Raosch
Germany, Rheinische Lehr- und Versuchsanstalt für Bienenzucht, Mayen-Eifel: G. Götze.
Germany, Berlin-Zehlendorf: L. Armbruster.
India, Punjab Agricultural College, Lyallpur: S. Singh.
Italy, Instituto Zoologico della R. Università di Bologna: Anita Vecchi.
Switzerland, Verein Deutsch-Schweizerischer Bienenfreunde, St. Gallen: M. Jüstrich.
Union of Soviet Socialist Republics, Ecological Laboratory, Moscow: W. W. Alpatov.
Union of Soviet Socialist Republics, Petrovka, Moscow: A. S. Michailoff.